You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000000312_00738
You are here: Home > Sequence: MGYG000000312_00738
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
TMHMM annotations
Basic Information help
| Species | Eisenbergiella sp900539715 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Firmicutes_A; Clostridia; Lachnospirales; Lachnospiraceae; Eisenbergiella; Eisenbergiella sp900539715 | |||||||||||
| CAZyme ID | MGYG000000312_00738 | |||||||||||
| CAZy Family | CBM32 | |||||||||||
| CAZyme Description | hypothetical protein | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 91547; End: 93877 Strand: + | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH20 | 432 | 769 | 1e-55 | 0.9673590504451038 |
| CBM32 | 76 | 187 | 6.7e-17 | 0.8306451612903226 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| cd06564 | GH20_DspB_LnbB-like | 3.19e-96 | 438 | 769 | 2 | 325 | Glycosyl hydrolase family 20 (GH20) catalytic domain of dispersin B (DspB), lacto-N-biosidase (LnbB) and related proteins. Dispersin B is a soluble beta-N-acetylglucosamidase found in bacteria that hydrolyzes the beta-1,6-linkages of PGA (poly-beta-(1,6)-N-acetylglucosamine), a major component of the extracellular polysaccharide matrix. Lacto-N-biosidase hydrolyzes lacto-N-biose (LNB) type I oligosaccharides at the nonreducing terminus to produce lacto-N-biose as part of the GNB/LNB (galacto-N-biose/lacto-N-biose I) degradation pathway. The lacto-N-biosidase from Bifidobacterium bifidum has this GH20 domain, a carbohydrate binding module 32, and a bacterial immunoglobulin-like domain 2, as well as a YSIRK signal peptide and a G5 membrane anchor at the N and C termini, respectively. The GH20 hexosaminidases are thought to act via a catalytic mechanism in which the catalytic nucleophile is not provided by solvent or the enzyme, but by the substrate itself. |
| cd02742 | GH20_hexosaminidase | 1.45e-33 | 439 | 769 | 2 | 303 | Beta-N-acetylhexosaminidases of glycosyl hydrolase family 20 (GH20) catalyze the removal of beta-1,4-linked N-acetyl-D-hexosamine residues from the non-reducing ends of N-acetyl-beta-D-hexosaminides including N-acetylglucosides and N-acetylgalactosides. These enzymes are broadly distributed in microorganisms, plants and animals, and play roles in various key physiological and pathological processes. These processes include cell structural integrity, energy storage, cellular signaling, fertilization, pathogen defense, viral penetration, the development of carcinomas, inflammatory events and lysosomal storage disorders. The GH20 enzymes include the eukaryotic beta-N-acetylhexosaminidases A and B, the bacterial chitobiases, dispersin B, and lacto-N-biosidase. The GH20 hexosaminidases are thought to act via a catalytic mechanism in which the catalytic nucleophile is not provided by the solvent or the enzyme, but by the substrate itself. |
| pfam00728 | Glyco_hydro_20 | 4.01e-31 | 436 | 697 | 1 | 275 | Glycosyl hydrolase family 20, catalytic domain. This domain has a TIM barrel fold. |
| cd06562 | GH20_HexA_HexB-like | 2.28e-27 | 436 | 768 | 1 | 322 | Beta-N-acetylhexosaminidases catalyze the removal of beta-1,4-linked N-acetyl-D-hexosamine residues from the non-reducing ends of N-acetyl-beta-D-hexosaminides including N-acetylglucosides and N-acetylgalactosides. The hexA and hexB genes encode the alpha- and beta-subunits of the two major beta-N-acetylhexosaminidase isoenzymes, N-acetyl-beta-D-hexosaminidase A (HexA) and beta-N-acetylhexosaminidase B (HexB). Both the alpha and the beta catalytic subunits have a TIM-barrel fold and belong to the glycosyl hydrolase family 20 (GH20). The HexA enzyme is a heterodimer containing one alpha and one beta subunit while the HexB enzyme is a homodimer containing two beta-subunits. Hexosaminidase mutations cause an inability to properly hydrolyze certain sphingolipids which accumulate in lysosomes within the brain, resulting in the lipid storage disorders Tay-Sachs and Sandhoff. Mutations in the alpha subunit cause in a deficiency in the HexA enzyme and result in Tay-Sachs, mutations in the beta-subunit cause in a deficiency in both HexA and HexB enzymes and result in Sandhoff disease. In both disorders GM(2) gangliosides accumulate in lysosomes. The GH20 hexosaminidases are thought to act via a catalytic mechanism in which the catalytic nucleophile is not provided by solvent or the enzyme, but by the substrate itself. |
| COG3525 | Chb | 9.51e-27 | 378 | 768 | 201 | 613 | N-acetyl-beta-hexosaminidase [Carbohydrate transport and metabolism]. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| AYE35153.1 | 7.45e-119 | 62 | 769 | 32 | 759 |
| QAS60556.1 | 7.45e-119 | 62 | 769 | 32 | 759 |
| AYE35146.1 | 1.95e-114 | 62 | 769 | 32 | 759 |
| QMI86523.1 | 3.67e-109 | 67 | 775 | 179 | 894 |
| QTU82680.1 | 2.14e-107 | 67 | 770 | 179 | 889 |
PDB Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| 6JQF_A | 4.78e-82 | 210 | 771 | 3 | 549 | Crystallizationanalysis of a beta-N-acetylhexosaminidase (Am2136) from Akkermansia muciniphila [Akkermansia muciniphila ATCC BAA-835] |
| 4H04_A | 8.79e-24 | 321 | 771 | 50 | 477 | Lacto-N-biosidasefrom Bifidobacterium bifidum [Bifidobacterium bifidum JCM 1254],4H04_B Lacto-N-biosidase from Bifidobacterium bifidum [Bifidobacterium bifidum JCM 1254],4JAW_A Crystal Structure of Lacto-N-Biosidase from Bifidobacterium bifidum complexed with LNB-thiazoline [Bifidobacterium bifidum JCM 1254],4JAW_B Crystal Structure of Lacto-N-Biosidase from Bifidobacterium bifidum complexed with LNB-thiazoline [Bifidobacterium bifidum JCM 1254],5BXP_A LNBase in complex with LNB-LOGNAc [Bifidobacterium bifidum JCM 1254],5BXP_B LNBase in complex with LNB-LOGNAc [Bifidobacterium bifidum JCM 1254],5BXR_A LNBase in complex with LNB-NHAcDNJ [Bifidobacterium bifidum JCM 1254],5BXR_B LNBase in complex with LNB-NHAcDNJ [Bifidobacterium bifidum JCM 1254],5BXS_A LNBase in complex with LNB-NHAcCAS [Bifidobacterium bifidum JCM 1254],5BXS_B LNBase in complex with LNB-NHAcCAS [Bifidobacterium bifidum JCM 1254],5BXT_A LNBase in complex with LNB-NHAcAUS [Bifidobacterium bifidum JCM 1254],5BXT_B LNBase in complex with LNB-NHAcAUS [Bifidobacterium bifidum JCM 1254] |
| 6YHH_A | 4.84e-22 | 383 | 772 | 92 | 480 | X-rayStructure of Flavobacterium johnsoniae chitobiase (FjGH20) [Flavobacterium johnsoniae UW101],6YHH_B X-ray Structure of Flavobacterium johnsoniae chitobiase (FjGH20) [Flavobacterium johnsoniae UW101] |
| 4C7D_A | 1.59e-20 | 363 | 647 | 66 | 339 | Structureand activity of the GH20 beta-N-acetylhexosaminidase from Streptomyces coelicolor A3(2) [Streptomyces coelicolor],4C7D_B Structure and activity of the GH20 beta-N-acetylhexosaminidase from Streptomyces coelicolor A3(2) [Streptomyces coelicolor],4C7F_A Structure and activity of the GH20 beta-N-acetylhexosaminidase from Streptomyces coelicolor A3(2) [Streptomyces coelicolor],4C7F_B Structure and activity of the GH20 beta-N-acetylhexosaminidase from Streptomyces coelicolor A3(2) [Streptomyces coelicolor] |
| 4C7G_A | 3.75e-20 | 363 | 647 | 66 | 339 | Structureand activity of the GH20 beta-N-acetylhexosaminidase from Streptomyces coelicolor A3(2) [Streptomyces coelicolor] |
Swiss-Prot Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| B2UPR7 | 5.77e-85 | 210 | 771 | 25 | 571 | Beta-hexosaminidase Amuc_2136 OS=Akkermansia muciniphila (strain ATCC BAA-835 / DSM 22959 / JCM 33894 / BCRC 81048 / CCUG 64013 / CIP 107961 / Muc) OX=349741 GN=Amuc_2136 PE=1 SV=1 |
| Q9HGI3 | 1.79e-18 | 319 | 695 | 71 | 456 | Beta-hexosaminidase OS=Emericella nidulans OX=162425 GN=nagA PE=1 SV=1 |
| P13723 | 1.83e-18 | 361 | 719 | 78 | 468 | Beta-hexosaminidase subunit A1 OS=Dictyostelium discoideum OX=44689 GN=hexa1 PE=1 SV=1 |
| B2UP57 | 2.65e-18 | 399 | 609 | 71 | 279 | Beta-hexosaminidase Amuc_2018 OS=Akkermansia muciniphila (strain ATCC BAA-835 / DSM 22959 / JCM 33894 / BCRC 81048 / CCUG 64013 / CIP 107961 / Muc) OX=349741 GN=Amuc_2018 PE=1 SV=1 |
| Q7WUL4 | 6.33e-18 | 381 | 768 | 80 | 448 | Beta-N-acetylhexosaminidase OS=Cellulomonas fimi OX=1708 GN=hex20 PE=1 SV=1 |
SignalP and Lipop Annotations help
This protein is predicted as SP
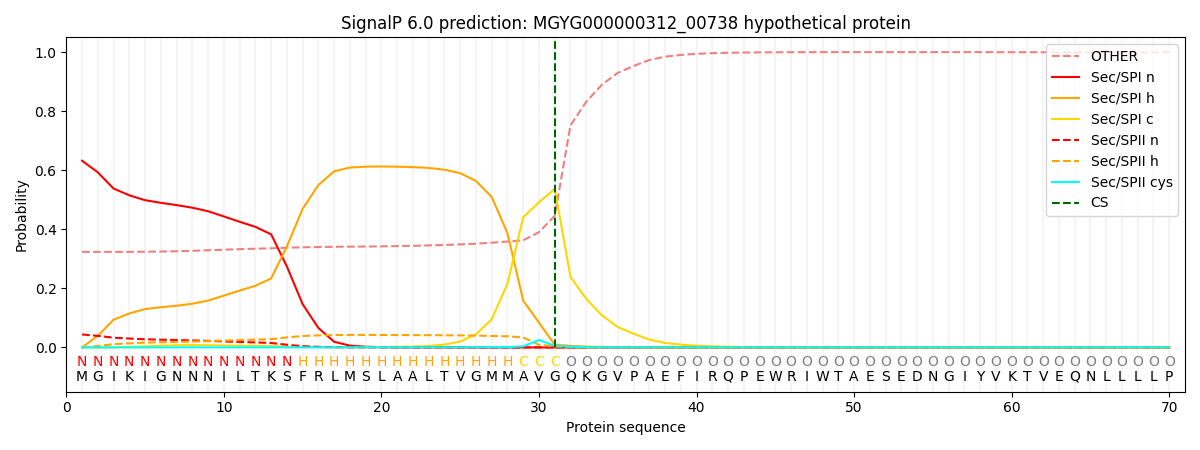
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.333032 | 0.620269 | 0.045328 | 0.000591 | 0.000389 | 0.000375 |