You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000003452_01581
You are here: Home > Sequence: MGYG000003452_01581
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
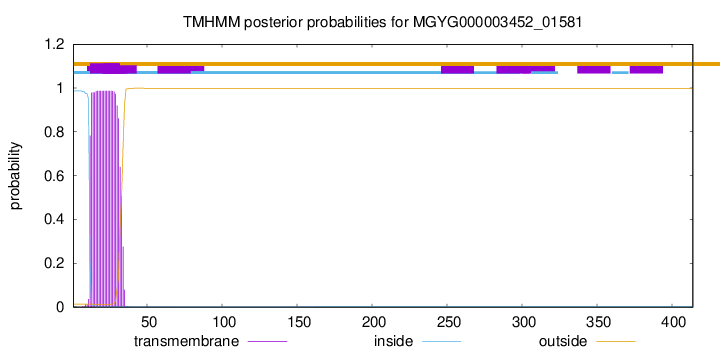
TMHMM annotations
Basic Information help
| Species | Bifidobacterium ruminantium | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Actinobacteriota; Actinomycetia; Actinomycetales; Bifidobacteriaceae; Bifidobacterium; Bifidobacterium ruminantium | |||||||||||
| CAZyme ID | MGYG000003452_01581 | |||||||||||
| CAZy Family | GH25 | |||||||||||
| CAZyme Description | hypothetical protein | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 78818; End: 80062 Strand: + | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH25 | 39 | 209 | 1.6e-31 | 0.9830508474576272 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| cd06417 | GH25_LysA-like | 1.32e-88 | 35 | 225 | 1 | 195 | LysA is a cell wall endolysin produced by Lactobacillus fermentum, which degrades bacterial cell walls by catalyzing the hydrolysis of 1,4-beta-linkages between N-acetylmuramic acid and N-acetyl-D-glucosamine residues. The N-terminal glycosyl hydrolase family 25 (GH25) domain of LysA has sequence similarity with other murein hydrolase catalytic domains while the C-terminal domain has sequence similarity with putative bacterial cell wall-binding SH3b domains. This domain family also includes LysL of Lactococcus lactis. |
| cd00599 | GH25_muramidase | 1.81e-27 | 37 | 218 | 2 | 186 | Endo-N-acetylmuramidases (muramidases) are lysozymes (also referred to as peptidoglycan hydrolases) that degrade bacterial cell walls by catalyzing the hydrolysis of 1,4-beta-linkages between N-acetylmuramic acid and N-acetyl-D-glucosamine residues. This family of muramidases contains a glycosyl hydrolase family 25 (GH25) catalytic domain and is found in bacteria, fungi, slime molds, round worms, protozoans and bacteriophages. The bacteriophage members are referred to as endolysins which are involved in lysing the host cell at the end of the replication cycle to allow release of mature phage particles. Endolysins are typically modular enzymes consisting of a catalytically active domain that hydrolyzes the peptidoglycan cell wall and a cell wall-binding domain that anchors the protein to the cell wall. Endolysins generally have narrow substrate specificities with either intra-species or intra-genus bacteriolytic activity. |
| pfam01183 | Glyco_hydro_25 | 8.70e-21 | 39 | 209 | 2 | 179 | Glycosyl hydrolases family 25. |
| cd06412 | GH25_CH-type | 1.38e-15 | 37 | 224 | 3 | 198 | CH-type (Chalaropsis-type) lysozymes represent one of four functionally-defined classes of peptidoglycan hydrolases (also referred to as endo-N-acetylmuramidases) that cleave bacterial cell wall peptidoglycans. CH-type lysozymes exhibit both lysozyme (acetylmuramidase) and diacetylmuramidase activity. The first member of this family to be described was a muramidase from the fungus Chalaropsis. However, a majority of the CH-type lysozymes are found in bacteriophages and Gram-positive bacteria such as Streptomyces and Clostridium. CH-type lysozymes have a single glycosyl hydrolase family 25 (GH25) domain with an unusual beta/alpha-barrel fold in which the last strand of the barrel is antiparallel to strands beta7 and beta1. Most CH-type lysozymes appear to lack the cell wall-binding domain found in other GH25 muramidases. |
| cd06415 | GH25_Cpl1-like | 6.50e-09 | 36 | 205 | 2 | 184 | Cpl-1 lysin (also known as Cpl-9 lysozyme / muramidase) is a bacterial cell wall endolysin encoded by the pneumococcal bacteriophage Cp-1, which cleaves the glycosidic N-acetylmuramoyl-(beta1,4)-N-acetylglucosamine bonds of the pneumococcal glycan chain, thus acting as an enzymatic antimicrobial agent (an enzybiotic) against streptococcal infections. Cpl-1 belongs to the CP family of lysozymes (CPL lysozymes) which includes the Cpl-7 lysin. Cpl-1 has a glycosyl hydrolase family 25 (GH25) catalytic domain with an irregular (beta/alpha)5-beta3 barrel and a C-terminal cell wall-anchoring module formed by six similar choline-binding repeats (ChBr's). The ChBr's facilitate the anchoring of Cpl-1 to the choline-containing teichoic acid of the pneumococcal cell wall. Other members of this domain family have an N-terminal CHAP (cysteine, histidine-dependent amidohydrolases/peptidases) domain similar to that of the firmicute CHAP lysins and associated with endopeptidase activity. The Cpl-7 lysin is also included here as is LysB of Lactococcus phage, and the Mur lysin of Lactobacillus phage. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| BAQ27398.1 | 8.12e-192 | 7 | 413 | 3 | 413 |
| VEG24063.1 | 8.12e-192 | 7 | 413 | 3 | 413 |
| ADB10077.1 | 8.12e-192 | 7 | 413 | 3 | 413 |
| AJE06518.1 | 1.24e-157 | 1 | 414 | 1 | 415 |
| AXR42563.1 | 2.97e-124 | 10 | 414 | 5 | 409 |
Swiss-Prot Hits help
SignalP and Lipop Annotations help
This protein is predicted as SP
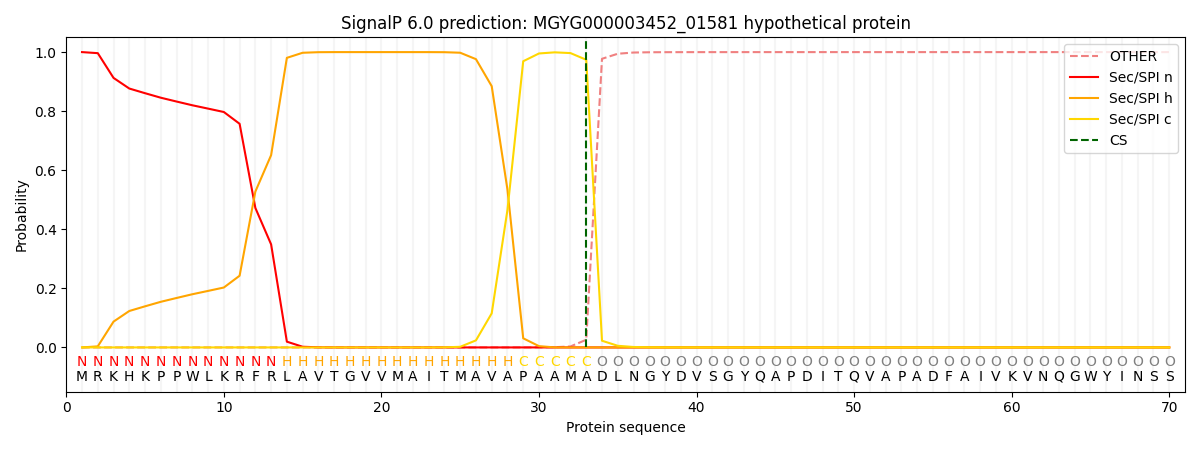
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.000310 | 0.998902 | 0.000199 | 0.000220 | 0.000189 | 0.000162 |