You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000004721_01528
You are here: Home > Sequence: MGYG000004721_01528
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
TMHMM annotations
Basic Information help
| Species | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Bacteroidota; Bacteroidia; Bacteroidales; Bacteroidaceae; Prevotella; | |||||||||||
| CAZyme ID | MGYG000004721_01528 | |||||||||||
| CAZy Family | GH31 | |||||||||||
| CAZyme Description | Alpha-xylosidase BoGH31A | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 4808; End: 7720 Strand: - | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH31 | 388 | 858 | 4.5e-136 | 0.9976580796252927 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| pfam01055 | Glyco_hydro_31 | 8.22e-169 | 389 | 858 | 1 | 442 | Glycosyl hydrolases family 31. Glycosyl hydrolases are key enzymes of carbohydrate metabolism. Family 31 comprises of enzymes that are, or similar to, alpha- galactosidases. |
| cd06591 | GH31_xylosidase_XylS | 1.07e-151 | 408 | 753 | 1 | 322 | xylosidase XylS-like. XylS is a glycosyl hydrolase family 31 (GH31) alpha-xylosidase found in prokaryotes, eukaryotes, and archaea, that catalyzes the release of alpha-xylose from the non-reducing terminal side of the alpha-xyloside substrate. XylS has been characterized in Sulfolobus solfataricus where it hydrolyzes isoprimeverose, the p-nitrophenyl-beta derivative of isoprimeverose, and xyloglucan oligosaccharides, and has transxylosidic activity. All GH31 enzymes cleave a terminal carbohydrate moiety from a substrate that varies considerably in size, depending on the enzyme, and may be either a starch or a glycoprotein. The XylS family corresponds to subgroup 3 in the Ernst et al classification of GH31 enzymes. |
| COG1501 | YicI | 3.22e-147 | 379 | 946 | 227 | 759 | Alpha-glucosidase, glycosyl hydrolase family GH31 [Carbohydrate transport and metabolism]. |
| cd06598 | GH31_transferase_CtsZ | 2.81e-63 | 408 | 764 | 1 | 330 | CtsZ (cyclic tetrasaccharide-synthesizing enzyme Z)-like. CtsZ is a bacterial 6-alpha-glucosyltransferase, first identified in Arthrobacter globiformis, that produces cyclic tetrasaccharides together with a closely related enzyme CtsY. CtsZ and CtsY both have a glycosyl hydrolase family 31 (GH31) catalytic domain; CtsY belongs to a different subfamily. All GH31 enzymes cleave a terminal carbohydrate moiety from a substrate that varies considerably in size, depending on the enzyme, and may be either a starch or a glycoprotein. |
| cd06589 | GH31 | 4.78e-62 | 408 | 750 | 1 | 265 | glycosyl hydrolase family 31 (GH31). GH31 enzymes occur in prokaryotes, eukaryotes, and archaea with a wide range of hydrolytic activities, including alpha-glucosidase (glucoamylase and sucrase-isomaltase), alpha-xylosidase, 6-alpha-glucosyltransferase, 3-alpha-isomaltosyltransferase and alpha-1,4-glucan lyase. All GH31 enzymes cleave a terminal carbohydrate moiety from a substrate that varies considerably in size, depending on the enzyme, and may be either a starch or a glycoprotein. In most cases, the pyranose moiety recognized in subsite -1 of the substrate binding site is an alpha-D-glucose, though some GH31 family members show a preference for alpha-D-xylose. Several GH31 enzymes can accommodate both glucose and xylose and different levels of discrimination between the two have been observed. Most characterized GH31 enzymes are alpha-glucosidases. In mammals, GH31 members with alpha-glucosidase activity are implicated in at least three distinct biological processes. The lysosomal acid alpha-glucosidase (GAA) is essential for glycogen degradation and a deficiency or malfunction of this enzyme causes glycogen storage disease II, also known as Pompe disease. In the endoplasmic reticulum, alpha-glucosidase II catalyzes the second step in the N-linked oligosaccharide processing pathway that constitutes part of the quality control system for glycoprotein folding and maturation. The intestinal enzymes sucrase-isomaltase (SI) and maltase-glucoamylase (MGAM) play key roles in the final stage of carbohydrate digestion, making alpha-glucosidase inhibitors useful in the treatment of type 2 diabetes. GH31 alpha-glycosidases are retaining enzymes that cleave their substrates via an acid/base-catalyzed, double-displacement mechanism involving a covalent glycosyl-enzyme intermediate. Two aspartic acid residues have been identified as the catalytic nucleophile and the acid/base, respectively. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| BCS86029.1 | 0.0 | 20 | 970 | 18 | 981 |
| VEH16024.1 | 0.0 | 13 | 970 | 14 | 980 |
| QNT65144.1 | 0.0 | 8 | 970 | 3 | 983 |
| ADE81187.1 | 0.0 | 7 | 970 | 6 | 1007 |
| QVJ80354.1 | 0.0 | 7 | 970 | 6 | 1030 |
PDB Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| 5JOU_A | 0.0 | 21 | 970 | 22 | 955 | Bacteroidesovatus Xyloglucan PUL GH31 [Bacteroides ovatus],5JOV_A Bacteroides ovatus Xyloglucan PUL GH31 with bound 5FIdoF [Bacteroides ovatus] |
| 7KMP_A | 1.47e-290 | 40 | 968 | 71 | 975 | ChainA, Alpha-xylosidase [Xanthomonas citri pv. citri str. 306],7KNC_A Chain A, Alpha-xylosidase [Xanthomonas citri pv. citri str. 306] |
| 2XVG_A | 1.04e-265 | 44 | 969 | 64 | 985 | crystalstructure of alpha-xylosidase (GH31) from Cellvibrio japonicus [Cellvibrio japonicus],2XVK_A crystal structure of alpha-xylosidase (GH31) from Cellvibrio japonicus in complex with 5-fluoro-alpha-D-xylopyranosyl fluoride [Cellvibrio japonicus],2XVL_A crystal structure of alpha-xylosidase (GH31) from Cellvibrio japonicus in complex with Pentaerythritol propoxylate (5 4 PO OH) [Cellvibrio japonicus] |
| 6JR6_A | 2.69e-64 | 371 | 936 | 221 | 771 | Flavobacteriumjohnsoniae GH31 dextranase, FjDex31A [Flavobacterium johnsoniae UW101],6JR6_B Flavobacterium johnsoniae GH31 dextranase, FjDex31A [Flavobacterium johnsoniae UW101],6JR6_C Flavobacterium johnsoniae GH31 dextranase, FjDex31A [Flavobacterium johnsoniae UW101],6JR6_D Flavobacterium johnsoniae GH31 dextranase, FjDex31A [Flavobacterium johnsoniae UW101],6JR7_A Flavobacterium johnsoniae GH31 dextranase, FjDex31A, complexed with glucose [Flavobacterium johnsoniae UW101],6JR7_B Flavobacterium johnsoniae GH31 dextranase, FjDex31A, complexed with glucose [Flavobacterium johnsoniae UW101],6JR7_C Flavobacterium johnsoniae GH31 dextranase, FjDex31A, complexed with glucose [Flavobacterium johnsoniae UW101],6JR7_D Flavobacterium johnsoniae GH31 dextranase, FjDex31A, complexed with glucose [Flavobacterium johnsoniae UW101] |
| 6JR8_A | 3.09e-63 | 371 | 936 | 221 | 771 | Flavobacteriumjohnsoniae GH31 dextranase, FjDex31A, mutant D412A complexed with isomaltotriose [Flavobacterium johnsoniae UW101],6JR8_B Flavobacterium johnsoniae GH31 dextranase, FjDex31A, mutant D412A complexed with isomaltotriose [Flavobacterium johnsoniae UW101],6JR8_C Flavobacterium johnsoniae GH31 dextranase, FjDex31A, mutant D412A complexed with isomaltotriose [Flavobacterium johnsoniae UW101],6JR8_D Flavobacterium johnsoniae GH31 dextranase, FjDex31A, mutant D412A complexed with isomaltotriose [Flavobacterium johnsoniae UW101] |
Swiss-Prot Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| A7LXT0 | 0.0 | 11 | 970 | 11 | 954 | Alpha-xylosidase BoGH31A OS=Bacteroides ovatus (strain ATCC 8483 / DSM 1896 / JCM 5824 / BCRC 10623 / CCUG 4943 / NCTC 11153) OX=411476 GN=BACOVA_02646 PE=1 SV=1 |
| Q9P999 | 8.79e-132 | 373 | 969 | 177 | 729 | Alpha-xylosidase OS=Saccharolobus solfataricus (strain ATCC 35092 / DSM 1617 / JCM 11322 / P2) OX=273057 GN=xylS PE=1 SV=1 |
| Q9F234 | 3.66e-58 | 386 | 904 | 230 | 721 | Alpha-glucosidase 2 OS=Bacillus thermoamyloliquefaciens OX=1425 PE=3 SV=1 |
| A2QTU5 | 1.93e-48 | 386 | 851 | 234 | 722 | Alpha-xylosidase A OS=Aspergillus niger (strain CBS 513.88 / FGSC A1513) OX=425011 GN=axlA PE=1 SV=1 |
| B3PEE6 | 7.46e-46 | 247 | 940 | 106 | 760 | Oligosaccharide 4-alpha-D-glucosyltransferase OS=Cellvibrio japonicus (strain Ueda107) OX=498211 GN=agd31B PE=1 SV=1 |
SignalP and Lipop Annotations help
This protein is predicted as SP
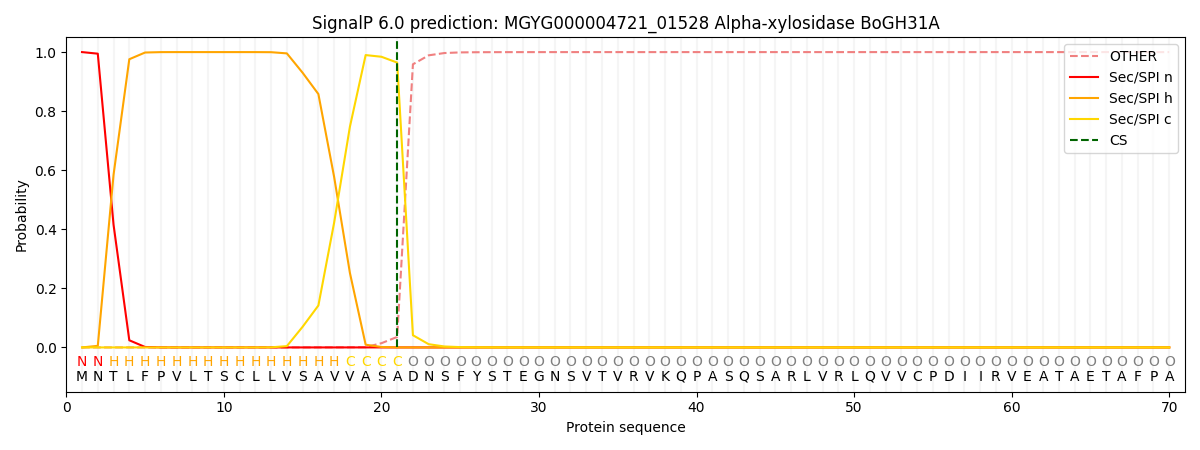
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.000210 | 0.999202 | 0.000177 | 0.000144 | 0.000140 | 0.000132 |